هنگامی که یک مولکول سیگنال دهنده به پروتئین گیرنده متصل می شود، باعث تغییر شکل آن می شود (تغییرات ساختاری/ ساختاری). لیگاندها (“اولین پیام رسان”) می توانند سیگنال ها را حمل کنند و با دیگر مولکول های درون سلولی، پیام رسان های دوم، تعامل داشته باشند. این سیگنال دهی داخل سلولی مولکول هایی به نام پیام رسان های دوم با وزن مولکولی کم خاص که عمر کوتاهی دارند و می توانند غلظتی را افزایش دهند (یا کاهش دهند) که باعث تغییر در عملکرد آنزیم ها یا پروتئین های غیر آنزیمی می شود. سیگنالینگ GPCR در این مقاله به تفصیل توضیح داده شده است.
برخی از نمونه های پیام رسان ثانویه هستند
- AMP 3،5 حلقوی (cAMP)،
- GMP 3،5 حلقوی (cGMP)،
- 1،2-دی آسیل گلیسرول (DAG)
- اینوزیتول 1،4،5-تری فسفات (IP3)
- تقریبا++ و فسفولیپیدهای مختلف اینوزیتول (فسفواینوزیتیدها).
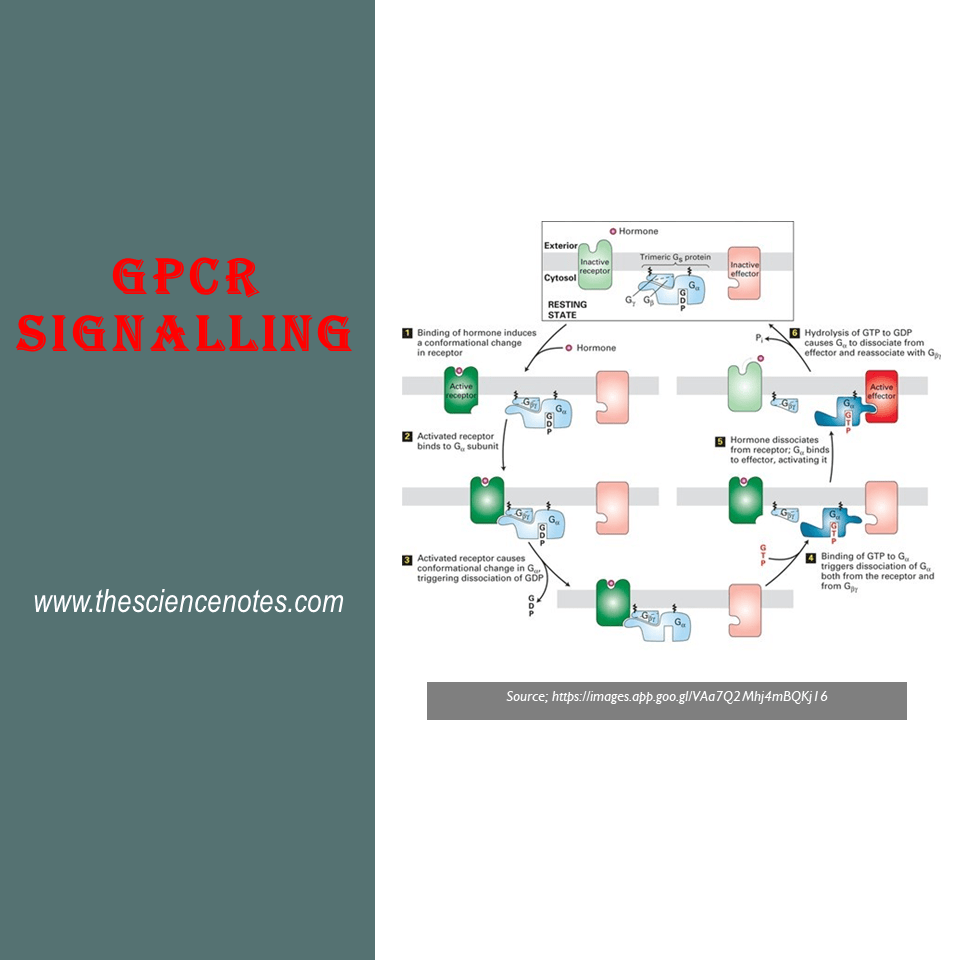
سیگنالینگ گیرنده جفت شده با پروتئین G
نحوه انتقال سیگنالها در مسیرهای مختلف درون سلولی که سیگنالها را در پایین دست گیرندههای سطح سلول فعال میکنند، متفاوت است. گیرنده های جفت شده با پروتئین G به طور غیر مستقیم آنزیم هایی را فعال می کند که پیام رسان های دوم درون سلولی را همانطور که در بالا ذکر شد تولید می کنند. آنها از طریق پروتئین های متصل شونده به GTP یا پروتئین های G برای انجام این کار ترکیب می شوند. به عنوان مثال – سیستم گیرنده آدرنرژیک که اپی نفرین (آدرنالین) را تشخیص می دهد. سیگنال دهی درون سلولی با گیرنده های جفت شده با پروتئین G (سیگنال دهی GPCR) شامل آبشارهای زیر است که در نکات زیر توضیح داده شده است.
گیرنده جفت شده 1.G-پروتئین در سیگنال دهی GPCR
GPCRها بزرگترین و متنوع ترین گروه گیرنده های غشایی هستند، زیرا ژنوم انسان حدود 800 GPCR مختلف را رمزگذاری می کند. لیگاندهای مختلف مسئول گیرندههای حسی مختلف از فوتونها، یونها، اسیدهای آمینه، رایحهها، طعمهای تلخ و شیرین، فرومونها، ایکوزانوئیدها، انتقالدهندههای عصبی، پپتیدها، پروتئینها و هورمونها با GPCRها که در نهایت طیف گستردهای از عملکردهای فیزیولوژیکی را تنظیم میکنند، شناخته شدهاند. .
ساختار یک GPCR
اگرچه GPCR ها دارای انواع مولکول های سیگنالینگ هستند، اما معماری مشترکی دارند که در طول تکامل حفظ شده است. آنها به عنوان شناخته می شوند گیرنده های هپتا مارپیچی، گیرنده های سرپانتین یا هفت گیرنده گذرنده به دلیل یک بسته هفت مارپیچ از GPCR ها که انعطاف پذیر است و می تواند بسیاری از ترکیبات را تغییر دهد. با توجه به سیگنال ها یا لیگاندهای دریافتی، آنها می توانند ترکیبات مختلف را تثبیت کنند و بنابراین سیگنال را به اعضای مختلف خانواده پروتئین هتروتریمری G منتقل کنند.
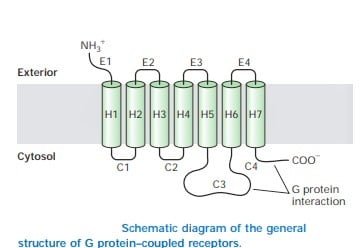
منبع: Lodish, Harvey F. Molecular Cell Biology. نیویورک: WH Freeman and Co، ویرایش پنجم
بنابراین، تعامل به طور عملکردی با گیرنده های پوشاننده غشاء و عوامل موثر انجام می شود.
2. پروتئین های سوئیچ GTPase
دو دسته از پروتئین های سوئیچ GTPase وجود دارد:
- پروتئین های G تریمریک (بزرگ) (پروتئین های گیرنده)
- پروتئین های مونومر (کوچک) G مانند Ras و پروتئین های مختلف Ras مانند.
پروتئین های G حاوی دامنه هایی مانند سوئیچ I و سوئیچ II هستند که فعالیت پروتئین های موثر خاص را از طریق برهمکنش مستقیم پروتئین-پروتئین زمانی که پروتئین G به GTP متصل می شود، کنترل می کنند. بنابراین، توانایی یک پروتئین G برای تعامل با پروتئین های دیگر و انتقال سیگنال در حالت “روشن” متصل به GTP و حالت “خاموش” وابسته به GDP متفاوت است.
(الف) همانطور که در شکل نشان داده شده است، دو دامنه به نام سوئیچ I (سبز) و سوئیچ II (آبی) در حالت فعال “روشن”، به فسفات پایانی GTP متصل می شوند. این از طریق فعل و انفعالات با گروههای آمید پایه یک باقیمانده ترئونین و گلیسین حفظ میشود.
(ب) آزاد شدن فسفات توسط هیدرولیز کاتالیز شده با GTPase باعث می شود سوئیچ I و سوئیچ II به یک ترکیب متفاوت شل شوند و حالت “خاموش” غیرفعال را امکان پذیر می کند.
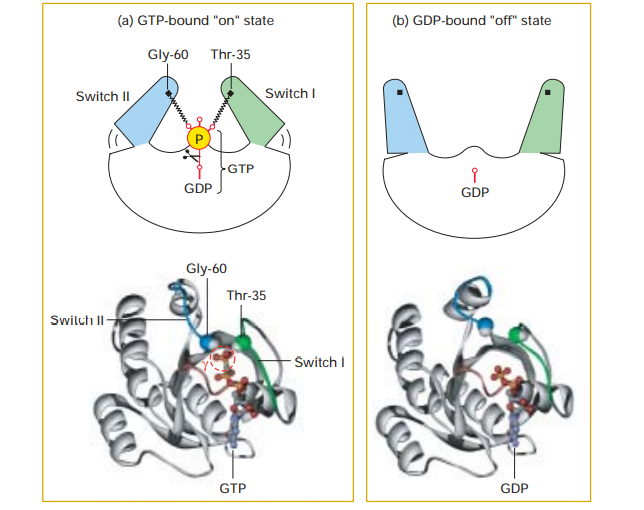
منبع: Lodish, Harvey F. Molecular Cell Biology. نیویورک: WH Freeman and Co، ویرایش پنجم
پروتئین های Trimeric G سیگنال های GPCR ها را در سیگنال دهی GPCR انتقال می دهند
آنها پروتئین های G نامیده می شوند زیرا نوکلئوتیدهای گوانین، GDP یا GTP را متصل می کنند. چندین خانواده از پروتئین های G تریمریک وجود دارد. پروتئین G هتروتریمری شامل سه زیر واحد پلی پپتیدی مختلف به نامهای α، β و γ است که توسط زنجیرههای لیپیدی که به صورت کووالانسی به زیر واحدهای α و γ متصل هستند، در غشای پلاسمایی نگهداری میشوند. پروتئین های متصل شونده به GTP هتروتریمر از طریق دنباله های آبگریز به غشاها متصل می شوند و به انتهای C زیر واحدهای آلفا و گاما متصل می مانند.
- محل اتصال نوکلئوتید گوانین در زیر واحد Gα وجود دارد. همانطور که در مکانیسم بالا ذکر شد، اتصال زیر واحد Gα به GTP آنها را “روشن” می کند، که منجر به یک آبشار انتقال سیگنال می شود، و هنگامی که به GDP متصل می شوند، “خاموش” می شوند و در نتیجه سیگنال دهی پایان می یابد.
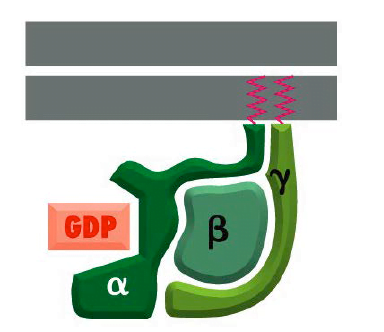
ساختار پروتئین G trimeric
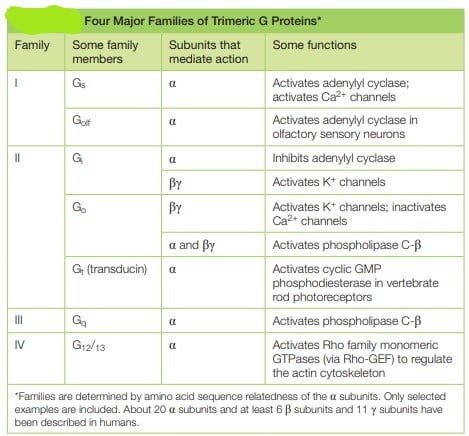
منبع; آلبرتز، بروس، الکساندر جانسون، جولیان لوئیس، مارتین راف، کیت رابرتز و پیتر والتر. 1381. زیست شناسی مولکولی سلول.
3. پروتئین کینازها و فسفاتازها در سیگنال دهی GPCR
فعال شدن تمام گیرنده های سطح سلول در نهایت منجر به تغییر در فسفوریلاسیون پروتئین از طریق فعال شدن پروتئین کینازها یا پروتئین فسفاتازها از طریق واسطه های ثانویه می شود. ژنوم انسان قادر به رمزگذاری 500 پروتئین کیناز و 100 فسفاتاز مختلف است. دو نوع پروتئین کیناز در سلول های حیوانی وجود دارد: آنهایی که فسفات را به گروه هیدروکسیل باقی مانده های تیروزین اضافه می کنند و آنهایی که فسفات را به گروه هیدروکسیل باقی مانده های سرین یا ترئونین (یا هر دو) اضافه می کنند. با این حال، فسفاتازهایی که گروه های فسفات را حذف می کنند، می توانند با کینازها عمل کنند تا عملکرد پروتئین های مختلف را خاموش یا روشن کنند.
گیرنده خود دارای فعالیت ذاتی کیناز یا فسفاتاز در برخی از مسیرهای سیگنالینگ است، در حالی که در مسیرهای دیگر گیرنده با سیتوزولی یا کینازهای متصل به غشاء تعامل دارد.
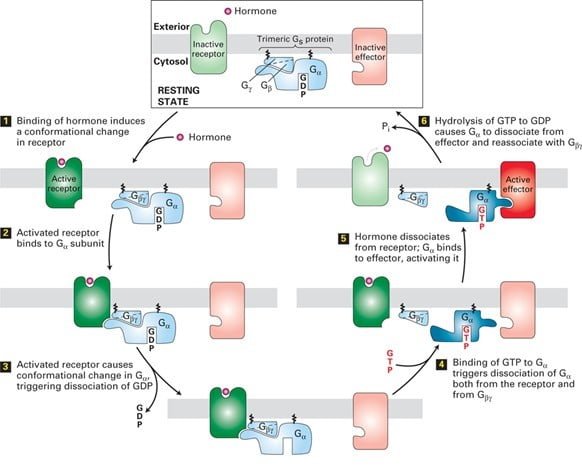
4. مکانیسم سیگنال دهی GPCR
سیگنال ها اغلب توسط G منتقل می شوندαیک پروتئین سوئیچ GTPase که به طور متناوب بین یک حالت فعال (“روشن”) با GTP محدود و یک حالت غیرفعال (“خاموش”) با تولید ناخالص داخلی. سایر زیرواحدهای ɤ و β که به هم متصل میمانند، گاهی سیگنالها را در سیگنالدهی GPCR انتقال میدهند.
یک مثال معمولی این است که گیرنده های اشغال شده با هورمون به عنوان GEF برای پروتئین های G عمل می کنند.
جدا کردن کاتالیزوری تولید ناخالص داخلی و اجازه دادن به GTP برای اتصال باعث تغییر در ساختار نواحی سوئیچ در G می شود و باعث جدا شدن آن از G می شود.α همانطور که در شکل نشان داده شده است، در هنگام اتصال لیگاند، با یک پروتئین مؤثر تعامل می کنند. هیدرولیز GTP سیگنالدهی را خاتمه میدهد و منجر به مونتاژ مجدد شکل trimeric میشود و سیستم را به حالت خاموش برمیگرداند. بنابراین، چرخه با اتصال یک مولکول لیگاند دیگر تکرار می شود. در برخی از مسیرها، پروتئین موثر توسط G آزاد فعال می شودɤβ زیر واحد
بیشتر بدانید
منابع
- ایواسا، جی.، مارشال، WF، و کارپ، جی. (2016). زیست شناسی سلولی و مولکولی کپور: مفاهیم و آزمایشات
- کراوس جی (2008). بیوشیمی انتقال و تنظیم سیگنال Wiley-VCH. صفحه 15. شابک 978-3527313976.
- آلبرتز، بروس، الکساندر جانسون، جولیان لوئیس، مارتین راف، کیت رابرتز و پیتر والتر. 1381. زیست شناسی مولکولی سلول. نیویورک: علم گارلند. ص 830-850
- لودیش، هاروی اف. زیست شناسی سلولی مولکولی. نیویورک: WH Freeman and Co، ویرایش پنجم. 543-567 ص


